энергии, требуют высокой скорости воспроизводства АТФ, обеспечить которую может только гликолиз, так как, в отличие от окисления, он протекает непосредственно в саркоплазме и не требует времени на доставку кислорода к митохондриям и доставку энергии от них к миофибриллам. Поэтому быстрые волокна предпочитают гликолитический путь воспроизводства АТФ и соответственно относятся к белым волокнам. За высокую скорость получения энергии белые волокна платят быстрой утомляемостью, так как гликолиз, как видно из уравнения реакции ведет к образованию молочной кислоты, накопление которой повышает кислотность среды и вызывает усталость мышцы и в конечном итоге останавливает ее работу.
Медленные волокна, характеризующиеся низкой активностью АТФазы, не требуют столь быстрого восполнения запасов АТФ и для обеспечения потребности в энергии используют путь окисления, то есть относятся к красным волокнам. Благодаря этому медленные волокна являются низко утомляемыми и способны поддерживать относительно небольшое, но длительное напряжение.
Существует и промежуточный тип волокон с высокой АТФазной активностью, и окислительно- гликолитическим путем воспроизводства АТФ.
Тип мышечного волокна зависит от мотонейрона его иннервирующего. Все волокна одного мотонейрона принадлежат к одному типу. Интересный факт – при подключении к быстрому волокну аксона медленного мотонейрона и наоборот, волокно перерождается, меняя свой тип. До недавнего времени существовало две точки зрения на зависимость типа волокна от типа мотонейрона, одни исследователи полагали, что свойства волокна определяются частотой импульсации мотонейрона, другие, что эффект влияния на тип волокна определяется гормоноподобными веществами поступающими из нерва (эти вещества до настоящего времени не выделены). Исследования последних лет показывают, что обе точки зрения имеют право на существование, воздействие мотонейрона, на волокно осуществляется обоими способами.
Как вы знаете на собственном опыте, человек обладает возможностью произвольно регулировать силу и скорость мышечного сокращения. Реализуется эта возможность несколькими способами. С одним из них вы уже знакомы – это управление частотой нервной импульсации. Подавая волокну одиночные команды на сокращение, можно добиться в нем легкого напряжения. Так, например, поддерживающие позу мышцы слегка напряжены, даже когда человек отдыхает. Повышая частоту импульсов можно увеличивать силу сокращения до максимально возможной для данного волокна в данных условиях работы, которая достигается при слиянии отдельных импульсов в тетанус.
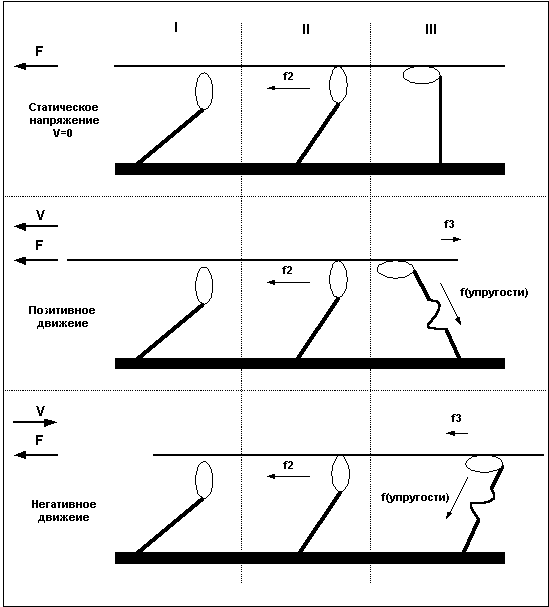
Сила, развиваемая волокном в состоянии тетануса, не всегда одинакова и зависит от характера и скорости движения. При статическом напряжении (когда длина волокна остается постоянной) сила, развиваемая волокном, больше чем при сокращении волокна, и чем быстрее сокращается волокно, тем меньшую силу оно способно развить. Максимальную же силу волокно развивает в условиях негативного движения, то есть при удлинении волокна.
При отсутствии внешней нагрузки волокно сокращается с максимальной скоростью. При увеличении нагрузки скорость сокращения волокна уменьшается и по достижении определенного уровня нагрузки падает до нуля, при дальнейшем увеличении нагрузки волокно удлиняется.
Причину в различии силы волокна при различных направлениях движения легко понять, рассмотрев уже приведенный ранее пример с гребцами и веслами. Дело в том, что после завершения «гребка» миозиновый мостик некоторое время находится в состоянии сцепления с нитью актина, представьте, что весло после гребка так же не сразу извлекается из воды, а находится погруженным еще некоторое время. В случае, когда гребцы плывут вперед (позитивное движение), весла, остающиеся погруженными в воду после завершения гребка, тормозят движение и мешают плыть, в то же время, если лодка буксируется назад, а гребцы сопротивляются этому движению, то погруженные весла так же мешают движению, и буксиру приходится прилагать большие усилия. То есть, при сокращении волокна, сцепленные мостики мешают движению и ослабляют силу волокна, при негативном движении – удлинении мышцы – не отцепленные мостики так же мешают движению, но в этом случае они как бы поддерживают опускающийся вес, что и позволяет волокну развивать большую силу. Легче всего понять различия между статическим напряжением, позитивным и негативным движением взглянув на Рис.5.
Итак, мы рассмотрели основные факторы, влияющие на силу и скорость сокращения отдельного волокна. Сила же сокращения целой мышцы зависит от количества волокон в данный момент времени вовлеченных в работу.
При поступлении от ЦНС (центральной нервной системы) к мотонейронам (расположенным в спинном мозге), возбуждающего сигнала (запускающий импульс), мембрана мотонейрона поляризуется, и он генерирует серию импульсов, направляемых по аксону к волокнам. Чем сильнее воздействие на мотонейрон (поляризация мембраны), тем выше частота генерируемого им импульса – от небольшой стартовой частоты (4-5 Гц), до максимально возможной, для данного мотонейрона, частоты (50 Гц и более). Быстрые мотонейроны способны генерировать гораздо более высокочастотный импульс, чем медленные, поэтому сила сокращения быстрых волокон гораздо больше подвержена частотной регуляции, чем сила медленных.
В то же время имеется и обратная связь с мышцей, от которой поступают тормозящие сигналы, уменьшающие поляризацию мембраны мотонейрона и снижающие его ответ.
Каждый мотонейрон имеет свой порог возбудимости. Если сумма возбуждающих и тормозящих сигналов, превышает этот порог и на мембране достигается необходимый уровень поляризации, то мотонейрон вовлекается в работу. Медленные мотонейроны имеют, как правило, низкий порог возбудимости, а быстрые – высокий. Мотонейроны же целой мышцы имеют широкий спектр значений этого параметра. Таким образом, при повышении силы сигнала ЦНС, активируется все большее число мотонейронов, а мотонейроны с низким порогом возбудимости увеличивают частоту генерируемого импульса.
Когда требуется легкое усилие, например, при ходьбе или беге трусцой, активируется небольшое число медленных мотонейронов и соответствующее число медленных волокон, ввиду высокой выносливости этих волокон такая работа может поддерживаться очень долго. По мере увеличения нагрузки ЦНС приходится посылать все более сильный сигнал, и большее число мотонейронов (а, следовательно, волокон), вовлекается в работу, а те, что уже работали, увеличивают силу сокращения, по причине увеличения частоты импульсации, поступающей от мотонейронов. По мере увеличения нагрузки в работу включаются быстрые окислительные волокна, а по достижения определенного уровня нагрузки (20%-25% от максимума), например, во время подъема в гору или финального спурта, силы окислительных волокон становится недостаточно, и посылаемый ЦНС сигнал включает в работу быстрые – гликолитические волокна. Быстрые волокна значительно повышают силу сокращения мышцы, но, в свою очередь, быстро утомляются, и в работу вовлекается все большее их количество. Если уровень внешней нагрузки не уменьшается, работу в скором времени приходится останавливать из-за усталости, в результате накопления молочной кислоты.
Скорость движения мышцы зависит от соотношения внешней нагрузки и количества вовлеченных в движение волокон. Мобилизация относительно небольшого количества волокон, приведет к медленному движению, как только совместная сила сокращающихся волокон превысит уровень внешней силы. Мобилизация все большего количества волокон при той же внешней нагрузке приведет к увеличению скорости сокращения, а увеличение внешней нагрузки, или падение силы волокон в результате усталости, при неизменном количестве мобилизованных волокон, приведет к падению скорости сокращения.
При предельных нагрузках, например, при подъеме максимального веса или подъеме относительно небольшого веса, но с максимальной скоростью, сокращается сразу максимально возможное для данного индивида число волокон.
По вопросу вовлечения волокон при предельных нагрузках в литературе приводятся противоречивые сведения. Так ряд авторов утверждает, что даже у тренированных спортсменов при предельных нагрузках активно не более 85% от общего количества двигательных единиц в мышце. С другой