floor 13, so there is probably an emergency control center there, and so on. But how inadequate all this knowledge would be if you were never allowed to hear what was being said and all your inferences were based on watching the windows.
If you think this analogy is too gloomy, then remember that the most sophisticated MRI scanners do nothing more than show where the lights are on in the brain. The only thing they reveal is where there is increased blood flow at any given moment, and we infer from this that more neural activity is taking place there. But we are nowhere near being able to understand what is “said” in the brain. We have no idea how any specific concept, label, grammatical rule, color impression, orientation strategy, or gender association is actually coded.
When researching this book, I read quite a few latter-day arguments about the workings of the brain shortly after trawling through quite a few century-old discussions about the workings of biological heredity. And when these are read in close proximity, it is difficult not to be struck by a close parallel between them. What unites cognitive scientists at the turn of the twenty-first century and molecular biologists at the turn of the twentieth century is the profound ignorance about their object of investigation. Around 1900, heredity was a black box even for the greatest of scientists. The most they could do was make indirect inferences by comparing what “goes in” on one side (the properties of the parents) and what “comes out” on the other side (the properties of the progeny). The actual mechanisms in between were mysterious and unfathomable for them. How embarrassing it is for us, to whom life’s recipe has been laid bare, to read the agonized discussions of these giants and to think about the ludicrous experiments they had to conduct, such as cutting the tails off generations of mice to see if the injury would be inherited by the offspring.
A century later, we can see much further into the mechanisms of genetics, but we are still just as shortsighted in all that concerns the workings of the brain. We know what comes in on one side (for instance, photons into the eye), we know what goes out the other side (a hand pressing a button), but all the decision making in between still occurs behind closed doors. In the future, when the neural networks will have become as transparent as the structure of DNA, when scientists can listen in on the neurons and understand exactly what is said, our MRI scans will look just as sophisticated as cutting off mice’s tails.
Future scientists will not need to conduct primitive experiments such as asking people to press buttons while looking at screens. They will simply find the relevant brain circuits and see directly how concepts are formed and how perception, memory, associations, and any other aspects of thought are affected by the mother tongue. If their historians of ancient science ever bother to read this little book, how embarrassing it will seem to them. How hard it will be to imagine why we had to make do with vague indirect inferences, why we had to see through a glass darkly, when they can just see face-to-face.
But ye readers of posterity, forgive us our ignorances, as we forgive those who were ignorant before us. The mystery of heredity has been illuminated for us, but we have seen this great light only because our predecessors never tired of searching in the dark. So if you, O subsequent ones, ever deign to look down at us from your summit of effortless superiority, remember that you have only scaled it on the back of our efforts. For it is thankless to grope in the dark and tempting to rest until the light of understanding shines upon us. But if we are led into this temptation, your kingdom will never come.
APPENDIX
Humans can see light only at a narrow band of wavelength from 0.4 to 0.7 microns (thousandths of a millimeter), or, to be more precise, between around 380 and 750 nanometers (millionths of a millimeter). Light in these wavelengths is absorbed in the cells of the retina, the thin plate of nerve cells that line the inside of the eyeball. At the back of the retina there is a layer of photoreceptor cells that absorb the light and send neural signals that will eventually be translated into the color sensation in the brain.
When we look at the rainbow or at light coming out of a prism, our perception of color seems to change continuously as the wavelength changes (see figure 11). Ultraviolet light at wavelengths shorter than 380 nm is not visible to the eye, but as the wavelength starts to increase we begin to perceive shades of violet; from around 450 nm we begin to see blue, from around 500 green, from 570 yellow, from 590 orange shades, and then once the wavelength increases above 620 we see red, all the way up to somewhere below 750 nm, where our sensitivity stops and infrared light starts.
A “pure” light of uniform wavelength (rather than a combination of light sources in different wavelengths) is called monochromatic. It is natural to assume that whenever a source of light looks yellow to us, this is because it consists only of wavelengths around 580 nm, like the monochromatic yellow light of the rainbow. And it is equally natural to assume that when an object appears yellow to us, this must mean that it reflects light only of wavelengths around 580 nm and absorbs light in all other wavelengths. But both of these assumptions are entirely wrong. In fact, color vision is an illusion played on us by the nervous system and the brain. We do not need any light at wavelength 580 nm to perceive yellow. We can get an identical “yellow” sensation if pure red light at 620 nm and pure green light at 540 nm are superimposed in equal measures. In other words, our eyes cannot tell the difference between monochromatic yellow light and a combination of monochromatic red and green lights. Indeed, television screens manage to trick us to perceive any shade of the spectrum by using different combinations of just three monochromatic lights-red, green, and blue. Finally, objects that appear yellow to us very rarely reflect only light around 580 nm and more usually reflect green, red, and orange light as well as yellow. How can all this be explained?
Until the nineteenth century, scientists tried to understand this phenomenon of “color matching” through some physical properties of light itself. But in 1801 the English physicist Thomas Young suggested in a famous lecture that the explanation lies not in the properties of light but rather in the anatomy of the human eye. Young developed the “trichromatic” theory of vision: he argued that there are only three kinds of receptors in the eye, each particularly sensitive to light in a particular area of the spectrum. Our subjective sensation of continuous color is thus produced when the brain compares the responses from these three different types of receptors. Young’s theory was refined in the 1850s by James Clerk Maxwell and in the 1860s by Hermann von Helmholtz and is still the basis for what is known today about the functioning of the retina.
Color vision is based on three kinds of light-absorbing pigment molecules that are contained within cells of the retina called cones. These three types of cells are known as long-wave, middle-wave, and short-wave cones. The cones absorb photons and send on a signal about the number of photons they absorb per unit of time. The short- wave cones have their peak sensitivity around 425 nm-that is, on the border between violet and blue. This does not mean that these cones absorb photons only at 425 nm. As can be seen from the diagram below (and in color in figure 12), the short-wave cones absorb light at a range of wavelengths, from violet to blue and even some parts of green. But their sensitivity to light decreases as the wavelength moves away from the peak at 425 nm. So when monochromatic green light at 520 nm reaches the short-wave cones, a much smaller percentage of the photons are absorbed compared to light at 425 nm.
The second type of receptors, the middle-wave cones, have their peak sensitivity at yellowish green, around 530 nm. And again, they are sensitive (to a decreasing degree) to a range of wavelengths from blue to orange. Finally, the long-wave cones have their peak sensitivity quite close to the middle-wave cones, in greenish yellow, at 565 nm.
The cones themselves do not “know” what wavelength of light they are absorbing. Each cone by itself is color-blind. The only thing the cone registers is the overall intensity of light that it has absorbed. Thus, a short- wave cone cannot tell whether it is absorbing low-intensity violet light (at 440 nm) or high-intensity green light at (500 nm). And the middle-wave cone cannot tell the difference between light at 550 nm and light in the same intensity at 510 nm.
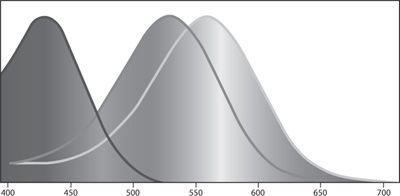
The (normalized) sensitivity of the short-wave, middle-wave, and long-wave cones as a function of wavelength.